GIP (Glucose-dependent insulinotropic polypeptide) regola il peso corporeo e il consumo di cibo mediante i neuroni GABAergici nei topi.

Contributo di Ragni
GIP (Glucose-dependent insulinotropic polypeptide) regola il peso corporeo e il consumo di cibo mediante i neuroni GABAergici nei topi.
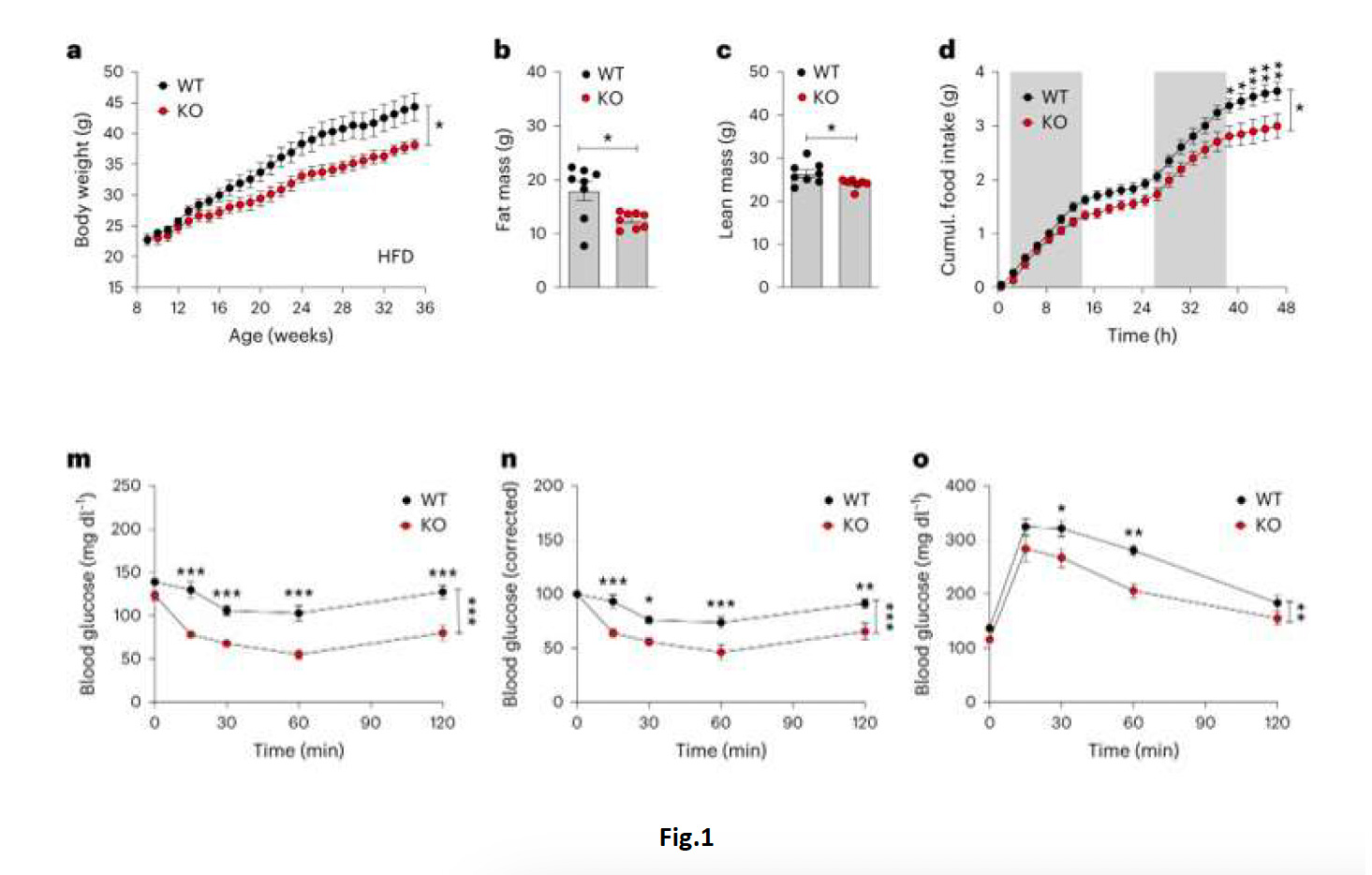
Lo sviluppo e utilizzo di agonisti per il recettore del glucagon-like peptide-1 (GLP-1) (GLP-1R) e per il recettore del Glucose-dependent insulinotropic polypeptide (GIP) (GIPR) hanno rappresentato un punto di svolta per la terapiadell’obesità e diabete; il co-agonismo GLP-1R- GIPR ha un’efficacia superiore, rispetto al solo agonismo GLP-1R,nell’indurre perdita di peso ma, nonostante ciò, il meccanismo d’azione di GIP rimane non chiarito. Numerose ricerche indicano che la capacità di degli agonisti GIPR di diminuire il peso corporeo (body weight – BW) risieda nella sua azioneanoressigenica centrale, ma le popolazioni e i mediatori neuronali coinvolti non sono stati ancora identificati. Nelmeccanismo d’azione dei co-agonisti GIPR/GLP1-R, l’agonismo per GIPR sembra inoltre essere indispensabile; adesempio, il co-agonista MAR709 ha una maggiore efficacia anoressigenica e anti-obesità rispetto al singolo acyl-GLP1 (un agonista GIPR a lunga durata d’azione), ma questa è abolita negli animali knockout per GIPR; gli agonisti GIPR attenuano l’effetto emetico dei GLP-1R, mentre anche l’effetto insulinotropico della tirzepatide (un doppio agonistaGLP-1R/GIPR) è attivato principalmente mediante l’agonismo del GIPR,(1–3). Questi dati quindi indicano come ilGIPR sia cruciale per l’effetto metabolico e per la tollerabilità dei co-agonisti GIPR/GLP1-R. Numerosi studi indicano che il GIPR è co-espresso, in diverse popolazioni cellulari dell’ipotalamo e del rombencefalo, con il trasportatoreSlc3a21 (o Vgat), un marker dei neuroni GABA, la cui azione è implicata nella regolazione dell’assunzione dei nutrienti(4). In questo lavoro, gli autori hanno perciò analizzato se l’effetto metabolico del co-agonismo GIPR/GLP1-Rdipenda dall’azione del GIPR nei neuroni GABAergici; a tale scopo, sono stati utilizzati topi knockout (KO) in cui il recettoreGIPR veniva soppresso esclusivamente nei neuroni esprimenti Vgat, cioè GABAergici. Gli animali KO, alimentati con dieta ad alto contenuto lipidico (high fat diet – HFD), mostravano un ridotto aumento di peso (fig. 1A) e consumo di cibo (foodintake – FI) (fig. 1D) rispetto ai topi wild-type (WT); I topi KO inoltre mostravano anche riduzione di massa grassa emagra (fig. 1B,1C), dei livelli plasmatici di glucosio e insulina e miglioramento della insulino-sensibilità (fig. 1 M,N,O).
Per quanto apparentemente controintuitivo, il dato è tuttavia confermato anche in altri lavori e fornisce il razionale, oltre cheper la sua attivazione, anche per l’antagonismo di GIPR nello sviluppo degli attuali farmaci antiobesità e antidiabetici (1),(5). Tuttavia, quando gli animali venivano alimentati con dieta standard (chow diet), sia il BW che il FI non differivano tra i gruppi KO e WT (fig. 2A, 2D), e ciò suggeriva quindi l’assenza di un ruolo per GIPR nella regolazionediretta sulla spesa energetica.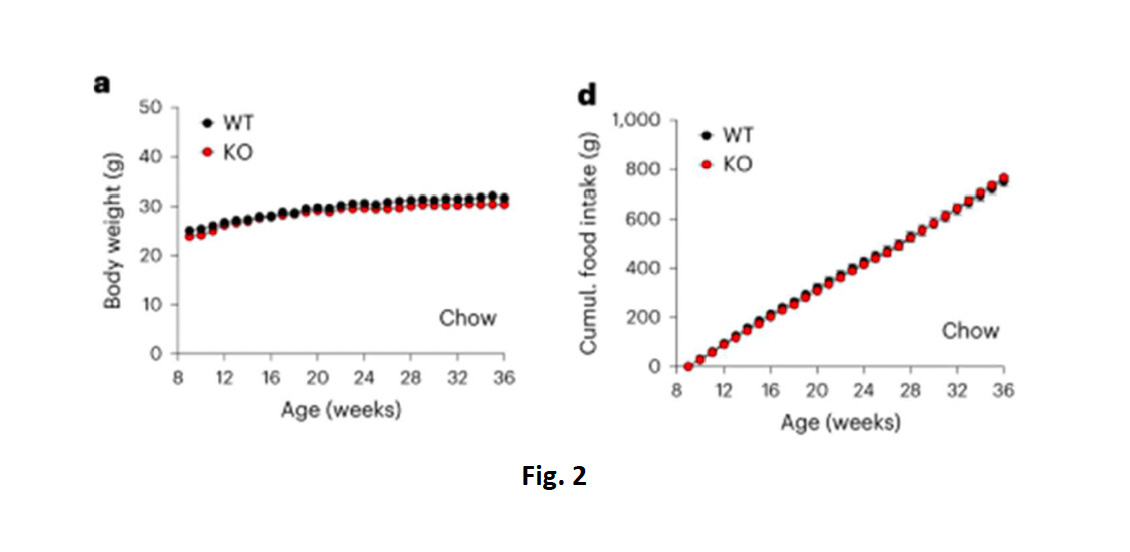
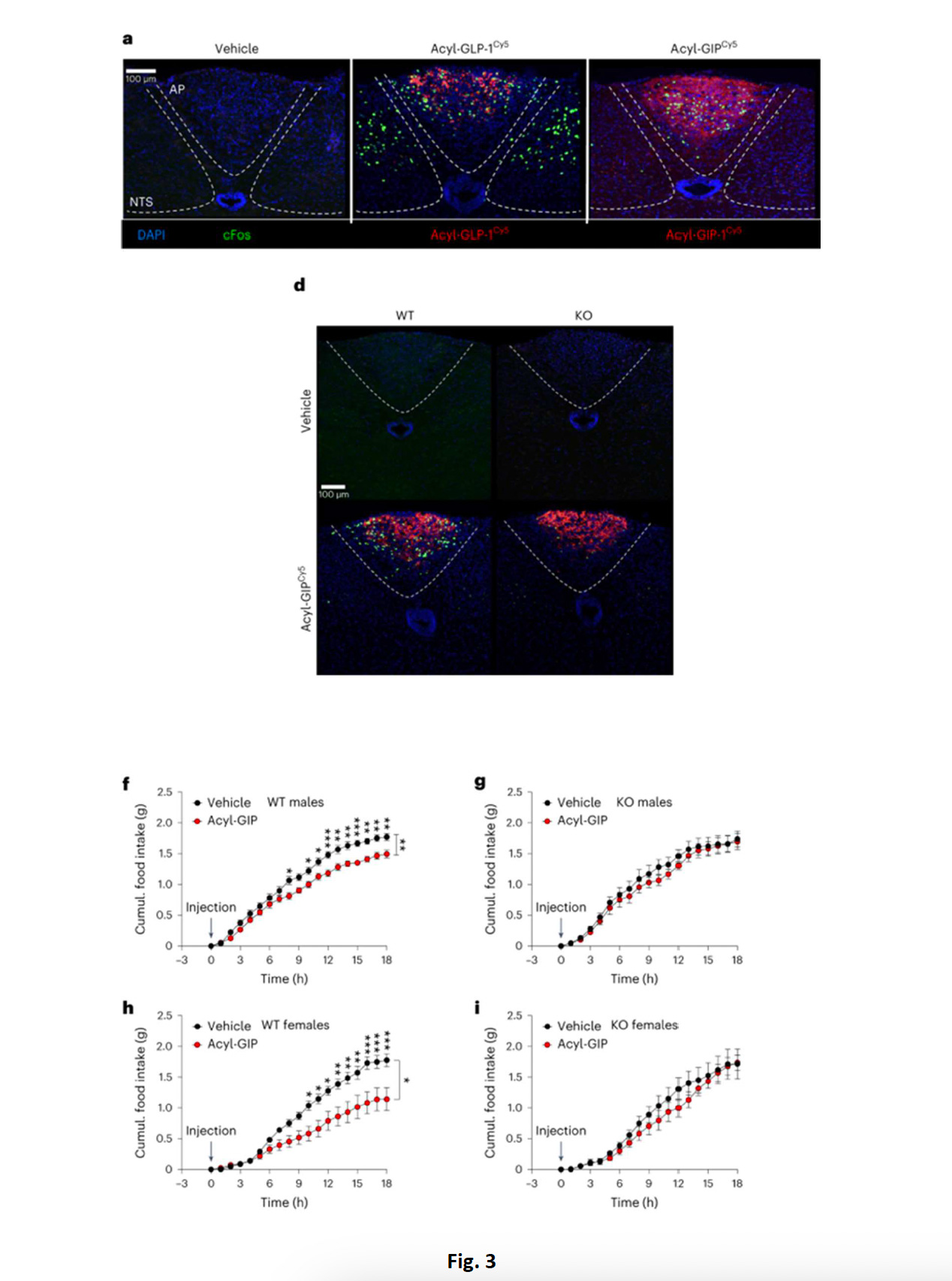
Allo scopo poi di verificare le conseguenze dell’assenza del GIPR nei neuroni GABAergici sull’agonismo di GLP- 1R eGIPR, I topi sono stati prima trattati in acuto con acyl-GLP1 e acyl-GIP, due agonisti a lunga durata d’azione, analizzando come marker di attività neuronale l’attività di c-fos; i due agonisti provocavano risposte diverse nei topi WT e KO; nei topiWT, acyl-GLP1 attivava c-fos sia nel nucleo del tratto solitario (NTS) che nell’area postrema (una regione particolarmente coinvolta nella regolazione del FI), mentre acyl-GIP solo in quest’ultima (fig. 3A), ma questa attivazione non si verificava nei topi Vgat-GIPR KO (fig. 3D) . Il dato perciò dimostra che GIP attiva c-fos nell’area postrema esclusivamente tramite i neuroni che esprimono Vgat, e che sono implicati nella trasmissione inibitoria GABAergica.Inoltre, mentre una singola iniezione di acyl-GIP diminuiva il FI in topi maschi e femmine WT alimentati in HFD, l’effetto eraassente nei topi KO (fig. 3F-I), indicando che il meccanismo anoressizzante dell’acyl-GIP richiede l’attivazione del GIPRlocalizzato nei neuroni inibitori GABAergici (Vgat-GIPR).
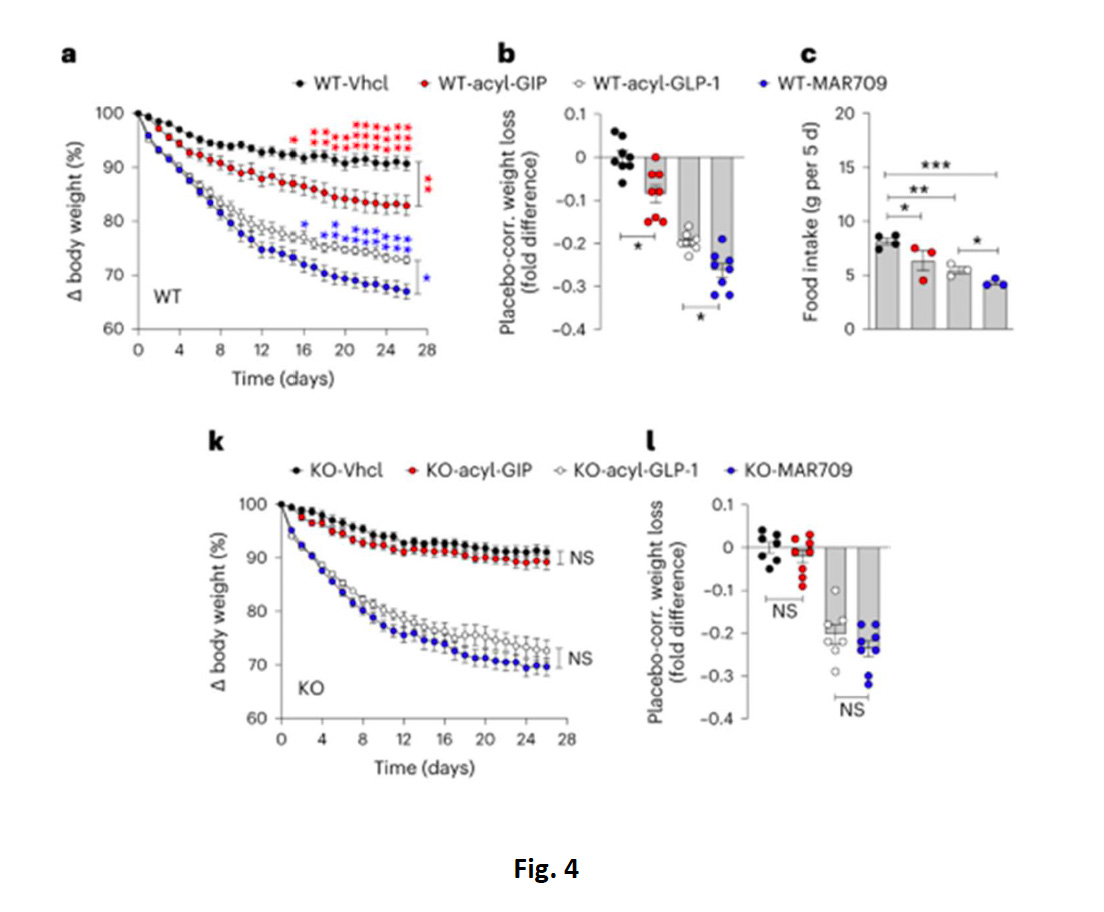
Successivamente, gli autori hanno verificato se il Vgat-GIPR fosse implicato anche nell’effetto degli agonisti GLP-1R eGIPR sul BW e parametri metabolici, trattando i topi alimentati con HFD in cronico (26 giorni) sia con acyl-GIP o acyl-GLP1che con il doppio agonista MAR709; come previsto, i topi WT trattati con acyl-GIP e acyl-GLP1 mostravano unadiminuzione del BW e FI rispetto ai controlli (fig. 4A-C), ma l’effetto era ancora una volta assente negli animali KO trattati con acyl-GIP, ma non con acyl-GLP1 (fig.4K, L). Inoltre, mentre nei topi WT sottoposti ad HFD, MAR709 induceva una perdita di peso e diminuzione di FI superiore a quella dei singoli acyl-GIP o acyl-GLP1, confermando quindi lamaggiore efficacia del doppio co-agonismo rispetto al singolo(fig. 4A-C), l’effetto superiore del MAR709 su BW e FI era tuttavia assente nei topi KO per Vgat-GIPR, dove la percentuale di perdita di peso era essenzialmente identica a quella indotta dal solo acyl-GLP1 (fig.4K, L) .
In conclusione, il lavoro dimostra che l’attività di GIPR nei neuroni GABAergici è indispensabile non solo per la capacità dell’acyl-GIP nel diminuire il FI e BW, ma anche per il maggiore effetto metabolico del co-agonista GLP-1R/GIPR MAR709 rispetto al solo acyl-GLP1
Commento:
Lo sviluppo dei nuovi agonisti GIPR/GLP-1R ha gettato le basi per una nuovo approccio farmacologico per la terapiadell’obesità; attualmente, sono diverse le strategie farmacodinamiche impiegate: insieme agli agonisti GLP-1 liraglutideand semaglutide, sono stati sviluppati sia singoli antagonisti GIPR, che doppi co- agonisti GIP/GLP1, come tirzepatide, o doppi agonisti GLP-1R/antagonisti GIPR, come AMG-133, con effetto superiore rispetto ai singoli. Il meccanismo d’azione alla base dell’effetto dimagrante/antidiabetico/anoressizzante delle incretine non è stato quindi ancora chiarito e, attualmente, sia l’antagonismo che l’agonismo per GIPR mostra effetti metabolici benefici (6) . Questo lavoro, nonostante confermi come il blocco dell’attività di GIPR sia efficace nel diminuire il BW e FI allo stesso modo della sua attivazione, dimostra come l’agonismo GIPR operi principalmente attraverso un meccanismo centrale, mediante ilcoinvolgimento dei neuroni GABAergici inibitori; inoltre, anche l’effetto superiore di un co- agonista come MAR709 è daattribuirsi all’azione del GIPR presente nei neuroni GABAergici. I risultati potrebbero quindi gettare le basi per lo sviluppo di nuove strategie farmacologiche e/o nuovi agonisti/antagonisti da accoppiare a quelle per GIPR/GLP-1R, estremamente versatili, già disponibili.
Referenze e ulteriori letture:
- Zhang Q, Delessa CT, Augustin R, Bakhti M, Colldén G, Drucker DJ, et al. The glucose-dependent insulinotropic polypeptide (GIP) regulates body weight and food intake via CNS-GIPR signaling. Cell 2021;33(4):833-844.e5.
- Borner T, Geisler CE, Fortin SM, Cosgrove R, Alsina-Fernandez J, Dogra M, et GIP Receptor Agonism Attenuates GLP-1 Receptor Agonist–Induced Nausea and Emesis in Preclinical Models. Diabetes.2021;70(11):2545–53.
- El K, Douros JD, Willard FS, Novikoff A, Sargsyan A, Perez-Tilve D, et The incretin co-agonist tirzepatiderequiresGIPRfor hormone secretion from human islets.Nat Metab. 2023;5(6):945–54.
- Jennings JH, Ung RL, Resendez SL, Stamatakis AM, Taylor JG, Huang J, et al. Visualizing hypothalamicnetwork dynamics for appetitive and consummatory Cell. 2015;160(3):516–27.
- Killion EA, Wang J, Yie J, Shi SDH, Bates D, Min X, et Anti-obesity effects of GIPR antagonists alone and in combination with GLP-1R agonists in preclinical models. Sci Transl Med. 2018;10(472).
- Campbell Targeting the GIPR for obesity: To agonize or antagonize? Potential mechanisms. Vol. 46,Molecular Metabolism. 2021.
Glucose-dependent insulinotropic polypeptide regulates body weight and food intake via GABAergic neurons in mice.
Liskiewicz A, Khalil A, Liskiewicz D, Novikoff A, Grandl G, Maity-Kumar G, Gutgesell RM, Bakhti M, Bastidas- PonceA, Czarnecki O, Makris K, Lickert H, Feuchtinger A, Tost M, Coupland C, Ständer L, Akindehin S, Prakash S, Abrar F, Castelino RL, He Y, Knerr PJ, Yang B, Hogendorf WFJ, Zhang S, Hofmann SM, Finan B, DiMarchi RD, Tschöp MH,Douros JD, Müller TD.
Nat Metab. 2023 Dec;5(12):2075-2085. doi: 10.1038/s42255-023-00931-7